【引文】
提高抗体对抗原亲和力是提高免疫分析方法灵敏度的有效手段,但是通过免疫动物产生的传统抗体,其亲和力容易受到免疫耐受的限制,且由于其结构复杂,不易于体外改造。相比而言,单域抗体(sdAb),也被称为纳米抗体,因其纳米级尺寸、良好的水溶性、对恶劣环境的高耐受性和易于基因操作的特点而更适合于体外诱导亲和成熟。目前已有研究者尝试了许多体外诱导纳米抗体亲和力提高的方法,但是突变的随机性使得筛选过程耗时费力,限制了这些方法的应用。本研究通过蛋白质同源建模和分子对接的组合策略以提供精准的蛋白质预测模型,选择纳米抗体抗赭曲霉毒素A(AOA-sdAb)作为模型抗体,分析赭曲霉毒素A(OTA)与AOA-sdAb间的分子相互作用,探索提高亲和力的可行途径。针对抗赭曲霉毒素A抗体中Nb28关键结合位点,通过丙氨酸扫描确认Gly53、Met79、Ser102和Leu149为关键氨基酸,通过对这4个关键氨基酸进行定点突变,构建了两个定点饱和突变文库,获得了亲和力提高的突变体Nb-G53Q&S102D,其半数最大抑制浓度(IC50)为0.29 ng/mL, KD值为52 nM,分别比原sdAb降低1.4倍和1.36倍。计算机模拟分析表明,氨基酸残基的氢键、疏水相互作用和侧链位阻对AOA-sdAb的结合亲和性至关重要。
【内容介绍】
1. 同源建模与分子对接
将Nb28的氨基酸序列提交至SWISSMODEL在线服务器获取其三维结构,从50个同源模板中选择相似度为54.88%,GMQE值为0.17,QMEAN评分为- 1.48的目标模板,其中z分数是通过将Nb28模型的归一化原始分数(综合QMEAN评分和单个平均力势项)与PDB数据库中的一组高分辨率x射线结构进行比较来计算的。如图1A所示,Nb28蛋白模型的Z-score位于一个相对准确的区域,表明Nb28模型的大部分氨基酸残基与预测特征吻合较好。此外,Ramachandran图分析显示,87个残基(94.6%)位于有利区域,5个残基(5.4%)位于可接受区域(图1B)。表明Nb28模型的所有骨架扭转角都在可信范围内。说明了所选择的Nb28同源性模型是合理的。
为了预测Nb28更稳定的三维结构,在隐式溶剂环境下对全原子分子进行了动态模拟。动力学仿真前200 ps的均方根偏差(RMSD)值如图1C所示。RMSD值在前10 ps快速增加,在10 – 200 ps范围内在1.2 Å左右轻微波动,具有轨迹的代表性结构特征,可认为预测的Nb28在200 ps内几乎是恒定的。利用Autodock 4.2将能量最小化后的稳定结构与OTA对接,如图1D所示,其中预测OTA与Nb28通过Gly53、Met79、Ser102、Val147和Leu149相互作用,涉及疏水相互作用和氢键。这些残基依次分布在Nb28的FR1、CDR1、CDR2和CDR3区域。这一结果说明Nb28的三个高变区中上述五个残基可能与其抗原结合活性有关。

图1. 最适模型的拟合曲线。(A)z得分结构数据库,(B)Nb28模型的Ramachandran图分析,(C)第一个200 ps Nb28的分子动力学模拟和结构,(D)Nb28和OTA的分子对接模型。
为了证实上述结论,进行了丙氨酸扫描实验,分别在Gly53、Met79、Ser102、Val147和Leu149位点引入了单点突变。然后,表达和纯化5个Nb28突变体(Nb-Gly53Ala、Nb-Met79Ala、NbSer102Ala、Nb-Val147Ala和Nb-Leu149Ala),并通过ELISA分析其活性。如图2A所示,与亲本sdAb Nb28相比,三个突变体(Nb-Gly53Ala、Nb-Met79Ala和Nb-Leu149Ala)的抗原结合活性显著降低。Nb-Ser102Ala和Nb-Val147Ala均保留了Nb28的抗原结合活性。然而,在间接竞争ELISA中,NbSer102Ala的敏感性比Nb28高1.5倍,NbVal147Ala没有观察到显著变化(图2B)。因此,Gly53、Met79、Ser102和Leu149可以被识别为Nb28与OTA相互作用的关键氨基酸位点,这证实了图1D中的分子对接结果。

图2. Nb28与丙氨酸扫描突变的活性分析。(A)Nb28及其突变体抗原结合能力间接ELISA标准曲线,(B)Nb28、Nb-Ser102Ala和Nb-Val147Ala的标准竞争抑制曲线。
2. 突变和噬菌体选择
使用Mut Express II Rapid Mutagenesis Kit V2将两个位点饱和突变引入Nb28的DNA序列,构建Gly53&Ser102-mLib和Met79&Leu149-mLib,计算存储容量。然后对两个突变文库进行4次生物筛选,从每个文库中随机抽取40个克隆进行噬菌体扩增,结果如图3A所示,所有Gly53&Ser102-mLib克隆均为阳性,随后,随机选取15个克隆进行DNA测序,得到6个不同的序列,如图3C所示。与噬菌体VHH28的序列相比,15个突变克隆的序列在Gly53(Nb-Gly53Asp、Nb-Gly53Lys、Nb-Gly53Leu和Nb-Gly53His)或Ser102(Nb-G53Q&S102D和Nb-Ser102Tyr)上均有两个位点突变。然而,从Met79和Leu149- mlib中选择的克隆在450 nm处的吸光度明显低于噬菌体VHH28(图3C),这表明引入Met79和Leu149位点的突变可以显著降低噬菌体VHH28的结合活性。

图3. 突变噬菌体克隆的选择和测序。(A)Gly53&Ser102-mLib和(B)Met79&Leu149-mLib噬菌体克隆的间接竞争性ELISA鉴定,(C)Gly53&Ser102-mLib克隆的6个DNA序列分析。
3. 突变体的表征和鉴定
6个不同序列突变体中VHH-G53D&S102Q、VHH-G53K&S102Q、VHH- G53Q & S102D三个克隆的灵敏度较高。选择IC50最小的克隆VHH-G53Q&S102D构建重组表达质粒。自动诱导表达和纯化后获得分子量约为18 kDa的Nb-G53Q&S102D,Nb-G53Q&S102D(IC50: 0.29±0.03 ng/mL)的灵敏度比预期的Nb28(IC50: 0.41±0.01 ng/mL)高1.41倍(图4B)。并对两种单克隆抗体的热稳定性和特异性进行了测试,结果表明Nb-G53Q&S102D在改变其亲本sdAbs中负责结合抗原的残基后仍保持了结构稳定性,具有更高的灵敏度、热稳定性和特异性。

图4. 突变体的鉴定和表征。(A)噬菌体克隆VHH28、VHH-G53D&S102Q、VHH-G53K&S102Q、VHH-G53L&S102Q、VHH-G53H&S102Q、VHH-G53Q&S102D、VHH-G53Q&S102Y的竞争抑制标准曲线,(B)Nb28和Nb-G53Q&S102D的竞争抑制标准曲线,(C)Nb28和Nb-G53Q&S102D分别在20、30、40、50、60、70、80和90°C孵育5分钟,在20°C下冷却10分钟,(D)分别在90°C下孵育0、5、15、25、35、45和55分钟,然后在相应时间的RT下冷却后的保留活性。
4. sdAb与OTA交互作用分析
采用“热泳与T跳”评价策略,进行了sdAb与OTA的相互作用亲和分析,NbG53Q&S102D的亲和力比Nb28高1.36倍,该结果表明,本研究中使用的策略在生成和筛选具有改进的OTA亲和力的sdAb方面是有效的。为了研究sdAb与OTA的作用机制,通过同源性建模和分子对接来探讨sdAb与OTA关键氨基酸的相互作用。与噬菌体克隆VHH28相比,从Met79和Leu149 - mlib中选择的突变噬菌体克隆的抗原结合活性显著降低(图3B)。此外,引入Nb28的Met79的丙氨酸突变可以使抗体失活(图2A)。这些结果可归因于Nb28的Met79被其他氨基酸取代,破坏了Met79与Tyr137之间形成的氢键(红点线),如图5B所示,对于突变的Nb-G53Q&S102D的第102位氨基酸,天冬氨酸和酪氨酸都可以与Lys107和OTA形成氢键,分别如图5C和D所示。但克隆VHH-G53Q&S102Y在噬菌体ELISA中的敏感性(IC50: 0.87±0.12 ng/mL)低于克隆VHH-G53Q&S102D(IC50: 0.11±0.01 ng/mL),推测S102Y侧链的位阻更强可能导致抗体亲和力降低。如图5E所示,Nb28的Leu149通过氢键和疏水相互作用与OTA相互作用。
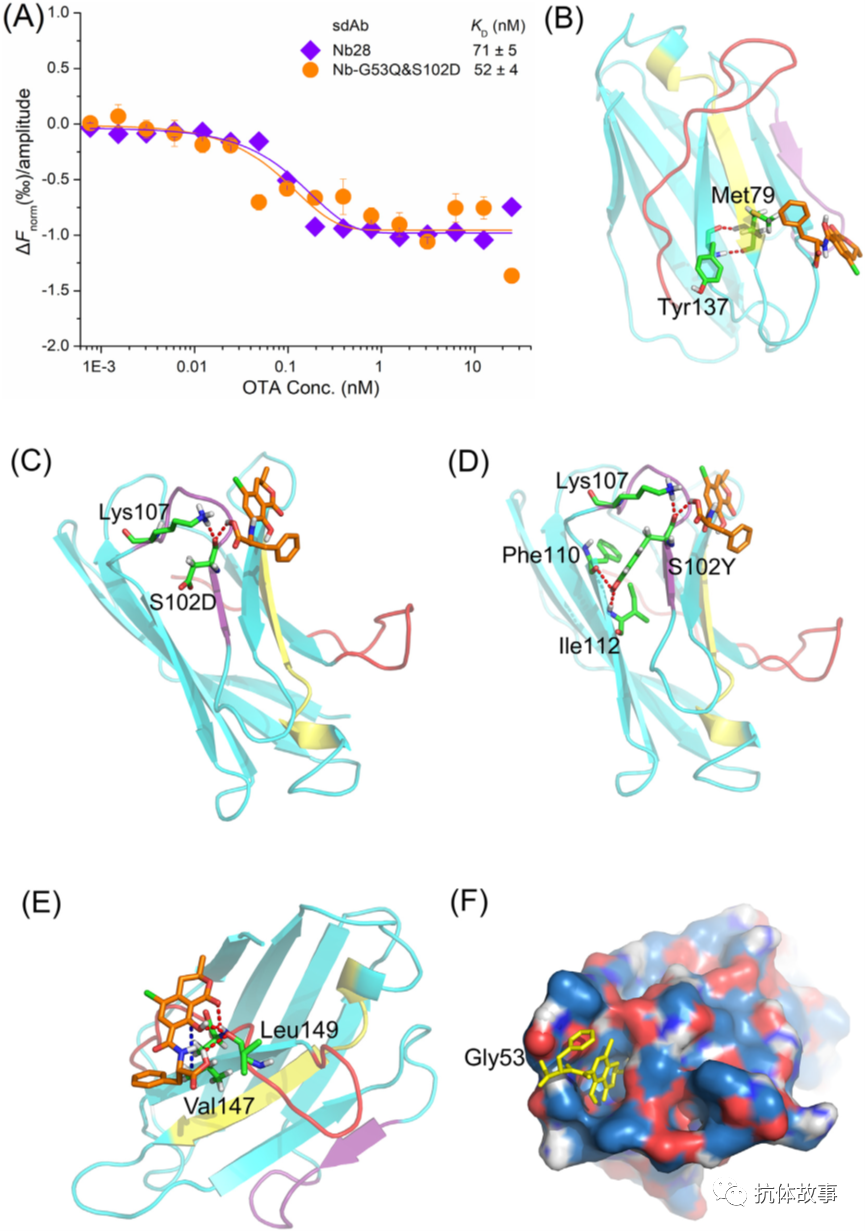
图5. sdAbs与OTA的相互作用分析。(A)Nb28和Nb-G53Q&S102D的归一化荧光变化,(B)Nb28的Met79与OTA的相互作用,(C)Nb-G53Q&S102D的S102D与OTA的相互作用,(D)Nb-G53Q&S102Y的S102Y与OTA的相互作用,(E)Nb28的Leu149与OTA的相互作用,(F)Nb28与OTA分子对接模型的结合孔。
虽然没有从Met79和Leu149- mlib中选择有效的噬菌体克隆(图3B),但这些相互作用力与Leu149的位阻可能对Nb28对OTA的结合活性和亲和力有重要影响。考虑到Met79在Nb28结合活性中的关键作用,引入Leu149的单点突变可能是提高抗体亲和力的另一种有效策略。如图4A所示,Gly53和Ser102对不同氨基酸的突变对VHH28的敏感性有很大影响。由于Q102和S102都是不带电的极性氨基酸,且具有相似的简单侧链,因此Nb活性口袋中涉及的第53位氨基酸的极性和侧链位阻可能是导致突变噬菌体克隆敏感性变化的主要原因(图5F)。随着G53D、G53K、G53L、G53H序列中第53位氨基酸侧链位阻的增加,相对突变噬菌体克隆的敏感性依次降低(图4A、图6)。VHH-G53D&S102Q、VHH-G53K&S102Q克隆的敏感性高于VHH28,这可能与G53D和G53K的极性有关。此外,第53位氨基酸的电荷可能对活性口袋有一定的影响,导致VHH-G53D&S102Q的IC50值低于VHH-G53K&S102Q(图4A)。

图6. 抗OTA sdAb 第53位4个氨基酸侧链位阻的比较。(A)G53D,(B)G53H,(C)G53L,(D)G53K。
尽管His53是极性氨基酸,但VHH-G53H&S102Q的灵敏度低于VHH28。这可能是由于组氨酸具有很强的侧链位阻作用。同样,亮氨酸的侧链位阻也可能是VHH-G53L&S102Q灵敏度低于VHH28的关键因素(图4A)。因此,通过修饰sdAb的关键氨基酸可以改变sdAb与OTA的相互作用。
【结论】
通过同源性建模、分子对接和丙氨酸扫描,鉴定出AOA-sdAb Nb28的4个关键氨基酸(Gly53、Met79、Ser102和Leu149)。对这些氨基酸进行配对,构建了两个位点饱和突变文库,从文库中筛选出了亲和力比亲本sdAb Nb28高1.36倍的突变Nb-G53Q&S102D。综上所述,同源建模和分子对接结合丙氨酸扫描和两点突变的策略适用于生成对OTA和其他毒性低分子量化合物具有更高亲和力的单克隆抗体,同时计算机模拟分析表明,氨基酸残基的氢键、疏水相互作用和侧链位阻对AOA-sdAb的结合亲和性至关重要。相对而言,使用同源建模和分子对接方法成本低、省时,为提高AOA-sdAb的结合亲和力提供了一种有效的策略,并为sdAb对其他低分子量化合物的结合提供了模型基础。
Wang, X. R., Chen, Q., Sun, Z. C., Wang, Y. D., Su, B. C., Zhang, C. H., Cao, H. M., & Liu, X. (2020). Nanobody affinity improvement: Directed evolution of the anti-ochratoxin A single domain antibody. International Journal of Biological Macromolecules,151, 312-321.
原文链接:https://doi.org/10.1016/j.ijbiomac.2020.02.180
本文来自于抗体故事,如您认为本已侵犯您的合法权益,请您及时联系我们删除!


